




 และยาทางเลือก")
 และวิธีจัดการ")





กลูคากอนเป็นฮอร์โมนเปปไทด์ที่ผลิตโดยเซลล์อัลฟาของตับอ่อน กลูคากอนช่วยเพิ่มความเข้มข้นของกลูโคสและกรดไขมันในกระแสเลือด และถือเป็นฮอร์โมน catabolic หลักของร่างกาย กลูคากอนยังใช้เป็นยารักษาโรคได้หลายอย่าง ผลของกลูคากอนตรงกันข้ามกับอินซูลินซึ่งช่วยลดระดับกลูโคสนอกเซลล์ Glucagon ผลิตจาก proglucagon ซึ่งเข้ารหัสโดยยีน GCG
ตับอ่อนจะหลั่งกลูคากอนเมื่อปริมาณกลูโคสในกระแสเลือดต่ำเกินไป กลูคากอนทำให้ตับมีส่วนร่วมในการสลายไกลโคเจน: เปลี่ยนไกลโคเจนที่เก็บไว้เป็นกลูโคสซึ่งถูกปล่อยออกสู่กระแสเลือด ระดับน้ำตาลในเลือดสูงกระตุ้นการหลั่งอินซูลิน อินซูลินช่วยให้กลูโคสถูกดูดซึมและนำไปใช้โดยเนื้อเยื่อที่ขึ้นกับอินซูลิน ดังนั้นกลูคากอนและอินซูลินจึงเป็นส่วนหนึ่งของระบบป้อนกลับที่ช่วยให้ระดับน้ำตาลในเลือดคงที่ กลูคากอนเพิ่มการใช้พลังงานและเพิ่มขึ้นภายใต้สภาวะความเครียด กลูคากอนเป็นฮอร์โมนในตระกูล secretin
หน้าที่ของกลูคากอน
โดยทั่วไปกลูคากอนจะช่วยเพิ่มความเข้มข้นของกลูโคสในเลือดโดยส่งเสริมการสร้างกลูโคเนซิสและไกลโคจีโนไลซิส กลูคากอนยังลดการสังเคราะห์กรดไขมันในเนื้อเยื่อไขมันและตับ กลูคากอนส่งเสริมการสลายไขมันในเนื้อเยื่อเหล่านี้ ซึ่งทำให้เกิดการปล่อยกรดไขมันเข้าสู่กระแสเลือด ซึ่งจะถูกแคแทบอลิซึมเพื่อสร้างพลังงานในเนื้อเยื่อ เช่น กล้ามเนื้อโครงร่าง เมื่อจำเป็น
กลูโคสถูกเก็บไว้ในตับในรูปแบบของพอลิแซ็กคาไรด์ไกลโคเจน ซึ่งเป็นกลูแคน (โพลีเมอร์ที่ประกอบด้วยโมเลกุลกลูโคส) เซลล์ตับ (hepatocytes) มีตัวรับกลูคากอน เมื่อกลูคากอนจับกับตัวรับกลูคากอน เซลล์ตับจะเปลี่ยนไกลโคเจนเป็นโมเลกุลกลูโคสแต่ละโมเลกุลและปล่อยออกสู่กระแสเลือด ในกระบวนการที่เรียกว่าไกลโคเจโนไลซิส เมื่อการจัดเก็บกลูโคสหมดลง กลูคากอนจะกระตุ้นให้ตับและไตสังเคราะห์กลูโคสเพิ่มเติมโดยการสร้างกลูโคนีเจเนซิส กลูคากอนปิดการไกลโคไลซิสในตับ ทำให้สารตัวกลางไกลโคไลติกถูกส่งไปยังกลูโคนีเจเนซิส
กลูคากอนยังควบคุมอัตราการผลิตกลูโคสผ่านการสลายไขมัน กลูคากอนกระตุ้นการสลายไขมันในมนุษย์ภายใต้สภาวะของการกดอินซูลิน (เช่น เบาหวานชนิดที่ 1)
การผลิตกลูคากอนดูเหมือนจะขึ้นอยู่กับระบบประสาทส่วนกลางผ่านวิถีทางที่ยังไม่ได้กำหนด ในสัตว์ที่ไม่มีกระดูกสันหลัง มีรายงานว่าการกำจัดก้านตามีผลต่อการผลิตกลูคากอน การตัดก้านตาในกุ้งตัวเล็กจะทำให้เกิดภาวะน้ำตาลในเลือดสูงที่เกิดจากกลูคากอน
กลไกการออกฤทธิ์ของกลูคากอน

กลูคากอนจับกับกลูคากอนรีเซพเตอร์ ซึ่งเป็นรีเซพเตอร์คู่โปรตีน G ซึ่งอยู่ในพลาสมาเมมเบรนของเซลล์ การเปลี่ยนแปลงโครงสร้างในตัวรับจะกระตุ้นโปรตีน G ซึ่งเป็นโปรตีนเฮเทอโรไตรเมอร์ที่มีหน่วยย่อย α, β และ γ เมื่อโปรตีน G มีปฏิสัมพันธ์กับตัวรับ มันจะผ่านการเปลี่ยนแปลงเชิงโครงสร้างที่ส่งผลให้เกิดการแทนที่โมเลกุลของ GDP ที่ถูกผูกไว้กับหน่วยย่อย α ด้วยโมเลกุล GTP การแทนที่นี้ส่งผลให้เกิดการปลดปล่อยหน่วยย่อย α จากหน่วยย่อย β และ γ หน่วยย่อยอัลฟ่ากระตุ้นเอนไซม์ตัวต่อไปในน้ำตก อะดีนิเลตไซคเลสโดยเฉพาะ
Adenylate cyclase ผลิต cyclic adenosine monophosphate (cyclic AMP หรือ cAMP) ซึ่งกระตุ้นโปรตีน kinase A (cAMP-dependent protein kinase) ในทางกลับกัน เอ็นไซม์นี้จะกระตุ้น phosphorylase kinase ซึ่งจากนั้น phosphorylates glycogen phosphorylase b (PYG b) จะแปลงเป็นรูปแบบแอคทีฟที่เรียกว่า phosphorylase a (PYG a) ฟอสฟอรีเลสเอเป็นเอนไซม์ที่ทำหน้าที่ปลดปล่อยกลูโคส 1-ฟอสเฟตจากไกลโคเจนโพลีเมอร์
ตัวอย่างของวิถีคือเมื่อกลูคากอนจับกับโปรตีนเมมเบรน โปรตีนเมมเบรนทำปฏิกิริยากับ Gɑβ𝛾 Gɑ แยกออกจากGβ𝛾และมีปฏิสัมพันธ์กับโปรตีน adenylyl cyclase ของเมมเบรน Adenylyl cyclase กระตุ้นการแปลง ATP เป็น cAMP แคมป์จับกับโปรตีนไคเนส A และฟอสโฟรีเลต phosphorylase kinase ที่ซับซ้อน ฟอสฟอรีเลต ฟอสโฟรีเลส ไคเนส ฟอสโฟรีเลต ฟอสโฟรีเลส ฟอสฟอรีเลตฟอสโฟรีเลสจับหน่วยกลูโคสจากไกลโคเจนเป็นกลูโคส 1-ฟอสเฟต
นอกจากนี้ การควบคุมการประสานงานของไกลโคไลซิสและกลูโคนีเจเนซิสในตับยังถูกปรับโดยสภาวะฟอสโฟรีเลชั่นของเอนไซม์ที่กระตุ้นการก่อตัวของไกลโคไลซิสที่มีศักยภาพที่เรียกว่าฟรุกโตส 2,6-บิสฟอสเฟต เอนไซม์โปรตีนไคเนส A (PKA) ที่ถูกกระตุ้นโดยน้ำตกที่สร้างโดยกลูคากอนจะทำให้ฟอสโฟรีเลตตกค้างซีรีนเดียวของสายพอลิเปปไทด์แบบไบฟังก์ชันที่มีทั้งเอนไซม์ฟรุกโตส 2,6-บิสฟอสฟาเตสและฟอสโฟฟรุกโตไคเนส-2 ฟอสโฟรีเลชั่นโควาเลนต์นี้เริ่มต้นโดยกลูคากอนกระตุ้นสารเดิมและยับยั้งสารหลัง สิ่งนี้ควบคุมปฏิกิริยาที่เร่งปฏิกิริยาฟรุกโตส 2,6-บิสฟอสเฟต (ตัวกระตุ้นที่มีศักยภาพของฟอสโฟฟรุกโตไคเนส-1 ซึ่งเป็นเอ็นไซม์ที่เป็นขั้นตอนหลักในการควบคุมไกลโคไลซิส) โดยการชะลออัตราการก่อตัว จึงยับยั้งการไหลของเส้นทางไกลโคไลซิสและทำให้เกิดกลูโคนีเจเนซิส เพื่อครอบงำ กระบวนการนี้สามารถย้อนกลับได้หากไม่มีกลูคากอน (และด้วยเหตุนี้การมีอินซูลิน)
การกระตุ้นกลูคากอนของ PKA ยังขัดขวางการทำงานของเอนไซม์ไกลโคไลติก ไพรูเวต ไคเนสในเซลล์ตับ
สรีรวิทยา
การผลิตกลูคากอน
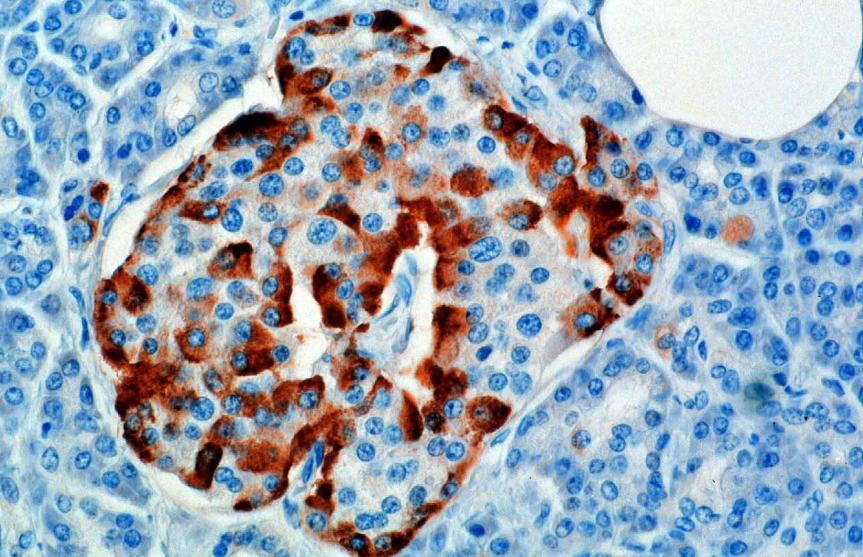
ฮอร์โมนถูกสังเคราะห์และหลั่งออกมาจากเซลล์อัลฟา (α-cells) ของเกาะ Langerhans ซึ่งอยู่ในส่วนต่อมไร้ท่อของตับอ่อน การผลิตซึ่งทำงานโดยอิสระถูกระงับ/ควบคุมโดยอะมิลิน ซึ่งเป็นฮอร์โมนเปปไทด์ที่หลั่งร่วมกับอินซูลินจากเซลล์ β ของตับอ่อน เมื่อระดับกลูโคสในพลาสมาลดลง การหลั่งอะมิลินที่ลดลงในเวลาต่อมาจะช่วยบรรเทาการปราบปรามของเซลล์ α ได้ ทำให้เกิดการหลั่งกลูคากอน
ในสัตว์ฟันแทะ เซลล์อัลฟาจะอยู่ที่ขอบด้านนอกของเกาะ โครงสร้างเกาะของมนุษย์มีการแบ่งแยกน้อยกว่ามาก และเซลล์อัลฟาจะกระจายไปทั่วเกาะในบริเวณใกล้เคียงกับเซลล์เบตา กลูคากอนยังผลิตโดยเซลล์อัลฟาในกระเพาะอาหาร
การวิจัยเมื่อเร็ว ๆ นี้แสดงให้เห็นว่าการผลิตกลูคากอนอาจเกิดขึ้นนอกตับอ่อนด้วย โดยลำไส้เป็นแหล่งที่มีแนวโน้มมากที่สุดของการสังเคราะห์กลูคากอนนอกตับอ่อน
ระเบียบกลูคากอน
การหลั่งของกลูคากอนถูกกระตุ้นโดย:
- ภาวะน้ำตาลในเลือดต่ำ
- อะดรีนาลีน (ผ่านทางตัวรับ adrenergic β2, α2 และ α1)
- อาร์จินีน
- อะลานีน (มักมาจากกล้ามเนื้อจากไพรูเวต/กลูตาเมตทรานส์อะมิเนชั่น
- อะเซทิลโคลีน
- Cholecystokinin
- โพลีเปปไทด์ยับยั้งกระเพาะอาหาร
การหลั่งของกลูคากอนถูกยับยั้งโดย:
- โซมาโตสแตติน
- อะมีลิน
- อินซูลิน (ผ่าน GABA)
- PPARγ/เรตินอยด์ X รีเซพเตอร์เฮเทอโรไดเมอร์
- เพิ่มกรดไขมันอิสระและกรดคีโตเข้าสู่กระแสเลือด
- เพิ่มการผลิตยูเรีย
- เปปไทด์คล้ายกลูคากอน-1
โครงสร้างกลูคากอน
กลูคากอนเป็นโพลีเปปไทด์กรดอะมิโน 29 ชนิด โครงสร้างหลักในมนุษย์คือ: NH2-His-Ser-Gln-Gly-Thr-Phe-Thr-Ser-Asp-Tyr-Ser-Lys-Tyr-Leu-Asp-Ser-Arg-Arg-Ala-Gln-Asp-Phe-Val-Gln-Trp -ลอย-เม็ท-อัศน-ธ-COOH.
โพลีเปปไทด์มีมวลโมเลกุล 3485 ดาลตัน กลูคากอนเป็นฮอร์โมนเปปไทด์ (ไม่ใช่สเตียรอยด์)
กลูคากอนถูกสร้างขึ้นจากความแตกแยกของโปรกลูคากอนโดยโปรโปรตีนคอนเวิร์เทส 2 ในเซลล์ α ของเกาะตับอ่อน ในเซลล์ L ของลำไส้ โปรกลูคากอนจะถูกแยกออกจากผลิตภัณฑ์อื่น glicentin, GLP-1 (an incretin), IP-2 และ GLP-2 (ส่งเสริมการเจริญเติบโตของลำไส้)
พยาธิวิทยา
ระดับกลูคากอนที่เพิ่มขึ้นอย่างผิดปกติอาจเกิดจากเนื้องอกในตับอ่อน เช่น กลูคาโกโนมา ซึ่งอาการต่างๆ ได้แก่ ภาวะเม็ดเลือดแดงเคลื่อนที่โดยเนโครไลติก กรดอะมิโนลดลง และน้ำตาลในเลือดสูง อาจเกิดขึ้นเพียงลำพังหรือในบริบทของเนื้องอกต่อมไร้ท่อหลายชนิด 1
กลูคากอนสูงเป็นสาเหตุหลักที่ทำให้เกิดภาวะน้ำตาลในเลือดสูงในผู้ป่วยเบาหวานชนิดที่ 1 ที่ไม่ได้รับการวินิจฉัยหรือรักษาได้ไม่ดี เมื่อเซลล์เบต้าหยุดทำงาน อินซูลินและ GABA ของตับอ่อนจะไม่มีอยู่อีกต่อไปเพื่อยับยั้งการผลิตกลูคากอนอิสระ เป็นผลให้กลูคากอนถูกปลดปล่อยออกจากเซลล์อัลฟาอย่างสูงสุด ทำให้เกิดการสลายตัวอย่างรวดเร็วของไกลโคเจนเป็นกลูโคสและการสร้างคีโตเจเนซิสอย่างรวดเร็ว พบว่ากลุ่มย่อยของผู้ใหญ่ที่เป็นเบาหวานชนิดที่ 1 ใช้เวลานานขึ้นโดยเฉลี่ย 4 เท่าในการเข้าถึงภาวะกรดซิตริกเมื่อให้โซมาโตสแตติน (ยับยั้งการผลิตกลูคากอน) โดยไม่มีอินซูลิน การยับยั้งกลูคากอนเป็นแนวคิดที่นิยมในการรักษาโรคเบาหวาน อย่างไรก็ตาม บางคนเตือนว่าการทำเช่นนี้จะทำให้เกิดโรคเบาหวานที่เปราะบางในผู้ป่วยที่มีระดับน้ำตาลในเลือดคงที่เพียงพอ
การไม่มีเซลล์อัลฟา (และด้วยเหตุนี้กลูคากอน) คิดว่าเป็นหนึ่งในอิทธิพลหลักในความผันผวนที่รุนแรงของระดับน้ำตาลในเลือดในการตั้งค่าของการผ่าตัดตับอ่อนทั้งหมด
ประวัติศาสตร์
ในปี ค.ศ. 1920 คิมบอลล์และเมอร์ลินศึกษาสารสกัดจากตับอ่อน และพบสารเพิ่มเติมที่มีคุณสมบัติระดับน้ำตาลในเลือดสูง พวกเขาอธิบายกลูคากอนในปี พ.ศ. 2466 ลำดับกรดอะมิโนของกลูคากอนอธิบายไว้ในช่วงปลายทศวรรษ 1950 ความเข้าใจอย่างถ่องแท้เกี่ยวกับบทบาทในด้านสรีรวิทยาและโรคยังไม่เกิดขึ้นจนกระทั่งทศวรรษ 1970 เมื่อมีการพัฒนาวิธีตรวจภูมิคุ้มกันด้วยรังสีเฉพาะ
.
Discussion about this post